|
|
АРИТМИЯ СЕРДЦА |
Нормальная и аномальная электрическая активность сердечных клеток


Потенциал покоя и потенциал действия в нормальных клетках синусового и атриовентрикулярного узлов
Электрическая активность клеток синусового и АВ-узлов весьма отличается от таковой в клетках специализированной проводящей системы желудочков или рабочего миокарда предсердий и желудочков, обсуждавшихся ранее. Благодаря своим необычным электрофизиологическим характеристикам клетки узлов часто принимают участие в инициации и поддержании аритмии. Ввиду существования значительных различий между клетками узла и другими клетками сердца их нормальные электрические характеристики целесообразно рассматривать отдельно.
Потенциал покоя
Клетки синусового узла обычно постоянно активны и редко находятся в покое, поэтому, строго говоря, при их описании не следует использовать термин «потенциал покоя». Однако максимальный диастолический потенциал (наиболее отрицательный уровень мембранного потенциала сразу после потенциала действия реполяризации) легко измеряется и оказывается значительно менее отрицательным (примерно на 20 мВ), чем максимальный диастолический потенциал волокон Пуркинье или клеток предсердий и желудочков (рис. 3.9).
Максимальный диастолический потенциал клеток АВ-узла по своей величине аналогичен наблюдаемому в клетках синусового узла. Значения внутриклеточной концентрации K+ (а значит, и величина ЕK) в клетках синусового узла, по-видимому, близки к зарегистрированным в сердечных клетках с гораздо более высоким потенциалом покоя [44]. Следовательно, более низкий мембранный потенциал клеток синусового и атриовентрикулярного узлов обусловлен более высоким отношением коэффициентов натриевой и калиевой проницаемости (РNа/РK) мембраны этих клеток по сравнению с клетками предсердий и желудочков или волокон Пуркинье. Правда, пока неясно, в какой степени более высокое отношение PNa—РК в клетках узлов обусловлено меньшей величиной РK, а в какой — большей величиной pna. Дальнейшие исследования, однако, покажут, имеют ли узловые клетки необычно высокую проницаемость для Na+ в покое или они обладают необычно низкой проницаемостью для K+.
Фазы деполяризации и реполяризации потенциала действия
В клетках синусового и атриовентрикулярного узлов скорость деполяризации в нулевую фазу намного ниже (1—20 В/с), чем в нормальных волокнах Пуркинье или клетках рабочего миокарда (см. рис. 3.9). Амплитуда потенциалов действия также весьма невелика (60—80 мВ); в некоторых клетках пик потенциала действия не превышает 0 мВ [3]. По сравнению с другими сердечными клетками более низкая скорость нарастания и меньшая амплитуда потенциала действия узловых клеток отражают значительно меньшую величину входящего тока перед нулевой фазой деполяризации в этих клетках. Имеющиеся в настоящее время данные однозначно свидетельствуют о том, что меньший по величине входящий ток в клетках синусового и атриовентрикулярного узлов течет не через быстрые натриевые каналы, а через медленные каналы и переносится ионами натрия и кальция (45— 47].
Такие потенциалы действия с нарастанием, зависящим от медленного входящего тока, часто называют «медленными ответами» в отличие от более обычных «быстрых ответов», нарастание в которых зависит от быстрого натриевого тока [20]. Из-за столь малой величины суммарного входящего тока и медленной деполяризации в нулевую фазу скорость проведения медленных потенциалов действия через узлы всегда низка (0,01— 0,1 м/с); именно такое медленное проведение в определенных условиях может обусловить возникновение нарушений ритма в тканях узлов. Как отмечалось ранее, медленные каналы для входящего тока имеют совершенно иные характеристики зависимости их воротного механизма от времени и потенциала по сравнению с быстрыми натриевыми каналами. Медленный входящий ток активируется и инактивируется значительно дольше, чем быстрый натриевый ток.
Поэтому после нарастания потенциала действия в узлах медленный входящий ток инактивируется только медленно, способствуя деполяризации мембраны в течение всей фазы плато потенциала действия. Активация зависимого от времени и потенциала выходящего калиевого тока вместе с инактивацией медленного входящего тока, по-видимому, вызывает реполяризацию клеток узлов, как это было описано для окончания потенциала действия в других сердечных клетках.
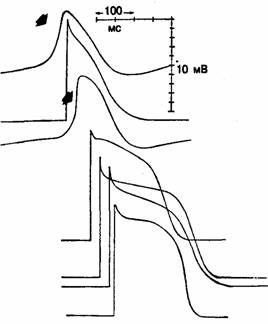
Рис. 3.9. Сравнение потенциалов действия синуса и АВ-узла (указаны стрелками) с потенциалами действия рабочего миокарда и волокон Пуркинье.
Запись потенциалов действия произведена в следующих областях сердца (начиная сверху): синусовый узел, предсердие, атриовентрикулярный узел, пучок Гиса, волокно Пуркинье в ложном сухожилии, терминальное волокно Пуркинье и рабочий миокард желудочков. Заметьте, что возрастание скорости и амплитуда потенциалов действия синуса и АВ-узла меньше аналогичных параметров в других клетках [3].
Проводимость медленных каналов для входящего тока восстанавливается после реполяризации мембраны также гораздо медленнее, чем проводимость быстрых натриевых каналов [20, 21]. В отличие от других сердечных клеток в узловых клетках при нанесении преждевременного стимула во время конечной фазы реполяризации не отмечается возникновения потенциала действия. В действительности достаточная инактивация проводимости медленных каналов для входящего тока может сохраняться даже после полной реполяризации в клетках, ставших абсолютно рефрактерными к стимуляции [48]. Реактивация происходит постепенно в течение всей диастолы; преждевременные импульсы, вызванные вскоре после полной реполяризации, имеют более медленное нарастание, меньшую амплитуду, чем нормальные импульсы, и распространяются медленнее.
Преждевременные импульсы, вызванные позднее во время диастолы, имеют соответственно более быстрое нарастание, более высокую амплитуду и, следовательно, проводятся быстрее [49]. Такое поведение отражает длительность процесса реактивации медленных каналов. Связанный с этим продолжительный рефрактерный период узловой ткани, а также значительное замедление проведения через нее преждевременных импульсов могут быть важными факторами инициации некоторых нарушений сердечного ритма.
Автоматизм
Клетки синусового узла обычно являются автоматическими, так что каждый последующий потенциал действия возникает в результате спонтанной диастолической деполяризации; клетки АВ-узла также способны возбуждаться автоматически, особенно если они не связаны с окружающим миокардом предсердий [50]. Это означает, по-видимому, что электротоническое взаимодействие клеток узла и предсердий подавляется автоматически через предсердно-узловое проведение. Автоматическая активность клеток синусового узла не может быть связана с тем же пейсмекерным током, который был ранее описан в волокнах Пуркинье. Изменения состояния воротного механизма мембранной проводимости, ответственной за нормальный автоматизм волокон Пуркинье, происходят только в диапазоне мембранных потенциалов от —90 до —60 мВ [36]. Такие изменения проводимости вряд ли способны объяснить развитие спонтанной диастолической деполяризации в клетках синусового узла, поскольку максимальный диастолический потенциал этих клеток обычно менее отрицателен, чем —60 мВ. Однако имеющиеся данные говорят о том, что пейсмекерный ток в синусовом узле переносится, по крайней мере частично, ионами К+ [38] и уменьшение этого выходящего тока при наличии постоянного фонового входящего тока приводит к постепенной деполяризации мембраны. Кроме того, важную роль играет, вероятно, входящий ток, который активируется при гиперполяризации и обозначается if [38].
|
|
|
К содержанию книги: АРИТМИИ СЕРДЦА. Механизмы, диагностика, лечение
|
Последние добавления:
Виноградский. МИКРОБИОЛОГИЯ ПОЧВЫ
Ферсман. Химия Земли и Космоса
Перельман. Биокосные системы Земли
Вильямс. Травопольная система земледелия