|
|
Биоминерализация в палеонтологии. Биоминералы. Причины скелетного минералообразования |
|
Биологическая минерализация - процесс образования живыми организмами неорганических соединений (минералов, минеральных агрегатов) и состоящих из них скелетных частей организмов. С общетеоретических позиций возникновение и эволюция биоминерализации представляет собой новый уровень интегрального взаимодействия живой и косной природы, когда живое не только использует косную материю в качестве среды и источника существования, но и создает "новое" косное вещество. Проблема биоминерализации носит комплексный общенаучный характер. В ней тесно переплетаются интересы и методы исследований молекулярной биологии, биохимии и биофизики, физиологии и медицины, физики твердого тела и геохимии. В течение последних 25-30 лет в этой области были проведены исследования в самых различных аспектах, опубликован ряд крупных коллективных трудов, регулярно проводятся региональные и международные совещания и симпозиумы [1335, 1337, 1398, 1560, 1943, 2291, 2301, 2374]. Особое значение в данной проблеме занимает палеонтология и комплекс геологических наук, так как возникновение, становление и развитие биоминерализации происходили в обозримом геологическом прошлом. Большинство объектов палеонтологии обязано своим происхождением биоминерализации, которая является выражением сложных метаболических процессов, происходящих в живых организмах. Таким образом, палеонтологические остатки как продукты биоминерализации, помимо сведений о морфологии и функциях, несут также информацию о существенно более тонких процессах (от молекулярных взаимодействий, приводивших к формированию биоминералов, до физиологических превращений на уровне клеток и тканей), благодаря которым образовывался скелет как морфологически и функционально неотъемлемая часть организма. Во многих своих аспектах этот уровень исследований ископаемых скелетных остатков по существу может рассматриваться как палеофизиологические исследования (см. 1.6.). Достигнутое понимание процесса формирования твердых скелетных частей организмов приводит к переоценке того понятия "скелет", которое традиционно сложилось в палеонтологии и закреплено в определениях, приводимых в "Палеонтологическом словаре" [799] и "Glossary of geology" [1068]. В этих формулировках недостаточно точно определен состав скелета и неполно отражены его функции. Не нашла в них отражения, в частности, морфообразующая функция скелета - одна из основных и наиболее важных, совершенно не обозначено физиологическое его значение.
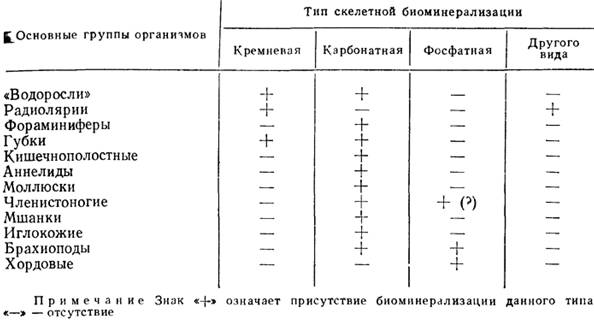
Достаточно распространенным является понимание скелета лишь как опорно-двигательной системы, при котором твердым образованиям большинства беспозвоночных вообще отказывается в праве называться скелетом. Эталоном самого понятия скелет, как и эталоном его изучения, признается лишь скелет позвоночных. Скелет позвоночных давно изучается в остеологии и палеонтологии не только с морфологических и сравнительно-анатомических, но и с гистологических и физиологических позиций. В частности, микроструктурные особенности как внутреннего, так и наружного скелета позвоночных, их гистологическая и физиологическая трактовка привели к развитию теорий образования костной ткани и т. д. (см. 2.4). Это приводит к определенному противопоставлению скелета позвоночных как более информативного во всех аспектах скелету беспозвоночных, что, несомненно, имеет некоторое объективное основание. Действительно, скелет позвоночных является в значительной степени клеточной тканью, т. е. тканью в гистологическом смысле; он, несомненно, является органом и даже системой органов с разнообразными, в том числе и физиологическими, функциями, обладает тесной интеграцией с "мягким телом". Твердые же образования беспозвоночных не являются клеточной тканью (не состоят из клеток и не вмещают клетки); они обычно не рассматриваются в качестве органа как части тела, состоящей из тканей и в значительной степени метаболически инертны. Однако достигнутые успехи в познании механизмов биоминерализации, показавшие большую общность образования минеральных составляющих как в скелете позвоночных, так и беспозвоночных, по крайней мере на уровне молекулярных взаимодействий при образовании самой минеральной фазы и при первичной минерализации, позволяют разрушить казалось бы существующую и существенную грань между скелетом позвоночных и беспозвоночных и подходить к изучению того и другого с методически адекватных позиций (см. 2.1). В дальнейшем целесообразно придерживаться самого широкого понятия "скелет" как плотной, твердой органической или минерализованной формообразующей части организма, которая может нести разнообразные функции, часто меняющиеся в онтогенезе и у филогенетически родственных форм. С палеонтологической точки зрения в настоящее время нецелесообразно рассматривать в качестве скелета филаментозные или тубулярные морфообразующие структуры, известные у многих одноклеточных (цитоплазматический скелет), хотя с общетеоретических позиций они являются не менее "настоящим" скелетом, чем скелет позвоночных. У некоторых простейших (например, радиолярий) строение минерализованного скелета скоррелировано и, вероятно, связано по происхождению со строением цитоплазматического скелета. Нельзя также считать скелетом твердые (иногда и минерализованные) структуры, которые не являются неотъемлемой частью организма и не являются морфообразующим субстратом собственно "мягкого тела" (домики ручейников, трубки некоторых полихет и т. д.), хотя они и могут обладать определенной степенью видоспецифичности. Иногда бывает трудно провести грань между собственно скелетом и домиком (у многих полихет) или между "продуктом" жизнедеятельности и скелетом (у строматолитов и многих "водорослей"). В дальнейшем мы будем касаться главным образом скелетной биоминерализации, хотя, несомненно, в сферу палеонтологии входят и аспекты нескелетного биоминералообразования. Известно около 40 минералов, которые образуются организмами. По степени распространенности и степени участия в скелетообразовании их можно разбить на четыре группы (табл. 1): 1) водный оксид кремния (аморфный кремнезем - опал); 2) карбонат кальция в трех минеральных видах - кальцит (в том числе высокомагнезиальный), арагонит, ватерит; 3) фосфаты кальция - апатит, франколит, даллит; 4) все прочие минералы.
Первое, что обращает на себя внимание, это связь типа биоминерализации с высотой организации организмов [55]. Кремневая биоминерализация преобладает у одноклеточных животных и растений и у параметазоа (губки). Во всех других случаях она не несет скелетной функции. Карбонатная биоминерализация распространена в большинстве таксономических групп, среди которых по количеству таксонов преобладают беспозвоночные. Среди одноклеточных карбонатная минерализация выполняет скелетную функцию у фораминифер и кокколитофорид; у позвоночных она ограничена и не несет скелетной функции. Фосфатная биоминерализация наиболее полно выражена у позвоночных, а среди беспозвоночных скелетообразующей является лишь у части брахиопод. Биоминералы четвертой группы представлены редко встречающимися минеральными видами, синтезируемыми отдельными группами организмов: минералы железа образуются некоторыми бактериями, целестин (сульфат стронция) продуцируется некоторыми радиоляриями и др. В большинстве случаев они не являются скелетообразующими. Эволюционные и физиологические причины скелетного минералообразования на основе кремния и биоминерализации четвертой группы неясны. Отчетливое же преобладание у живых организмов скелетов, образованных на основе кальциевых минералов, можно считать эволюционно детерминированным и выявить физиологические причины кальциевой биоминерализации. Кальций - один из наиболее биологически активных элементов, принимавший большое участие в достижении всех основных эволюционных уровней, последовательно пройденных живым веществом нашей планеты. Кальций необходим для нормального функционирования рибосом - аппарата трансляции генетического кода, он принимает непосредственное участие в полимеризации микротрубочек митотического веретена, широко известно значение кальция в специфическом слипании клеток многоклеточных животных в мускульном сокращении и в передаче нервных импульсов. Таким образом, участие кальция необходимо и для интеграции рибосом с клеткой, и для возникновения митоза, и для достижения уровня многоклеточности, возникновения мышечной и нервной систем. Как показано работами последних лет, Ca-связывающие протеины, выполняющие регулирующую и информационную роль в клетках, родственны по своей структуре и составу протеинам, входящим в минерально-органический комплекс скелетных тканей. Это доказывает, что возникновение скелетной биоминерализации на основе кальция является закономерной и необходимой ступенью, отражающей эволюцию метаболизма кальция и эволюцию организмов вообще. Представляется столь же закономерным переход с повышением уровня организации от карбонатной биоминерализации к минерализации с участием другого биологически активного элемента - фосфора. Различные аспекты эволюции процесса биоминерализации рассматривались в последние годы [54, 584, 1091]. Первичной физиологической функцией и причиной биоминерализации, по-видимому, была регуляция уровня содержания кальция в клетке и как необходимый результат этого - выведение кальция за пределы цитоплазмы. Минимальное требование для протекания этого процесса - наличие органических соединений, способных транспортировать ионы кальция, и соединений, способных связывать их в нерастворимый комплекс. Этот начальный этап биоминерализации (индуцируемый тип биоминерализации, по Г. Лоуэнштаму [1900]) моделируется образованием слизисто-карбонатных выделений, продуцируемых породообразователями строматолитов и многими другими группами "водорослей". Образование биоминералов может происходить внутриклеточно, внеклеточно или межклеточно. Биоминерализация индуцируемого типа часто не является скелетообразующей (строматолиты) и свойственна в основном низшим организмам. Но и у растений (Rodophyta, Siphonophyta, Charophyta) и, вероятно, у некоторых древнейших животных (Porifera, Archaeocyatha, Protozoa) она может быть основой скелетообразования. Интеграция процесса биоминерализации с образованием формообразующих, опорных и защитных структур дает начало минерализованному скелету. Более высокий уровень интеграции скелета и "мягкого тела" достигается, когда организм контролирует не только выведение кальция и его связывание в молекулярной и кристаллографической формах, но и объединение минеральных единиц в агрегаты определенной формы и структуры, а последних - в морфологически специфичную твердую часть организма - скелет ("матрицируемый" тип биоминерализации, по Г. Лоуэнштаму). Это приводит к появлению новых фракций органического матрикса, ответственных за формирование специфической морфологии минеральных единиц, за специфическую морфологию их агрегатов и в конечном итоге - за специфичность макроморфологии скелета. Другими словами, на этом этапе интеграции скелета и мягкого тела биоминерализация проявляется на нескольких уровнях организации скелетного вещества. Она может быть рассмотрена на уровне минералообразования, на уровне формирования минеральных агрегатов (т. е. на уровне скелетной ткани, на уровне формирования скелетных элементов, на уровне органов и системы органов), на уровне морфологии скелета как целого и как части организма. Эта организация в строении скелета организма, отражающая размерную соподчиненность, форму и взаимное расположение его внутренних элементов минеральных и органических, составляет структуру скелета. В известной степени операционалистский, феноменологический смысл имеет номенклатурное, терминологическое обозначение основных уровней организации скелетной ткани: макроструктурный, текстурный, микроструктурный, ультраструктурный и молекулярный [51, 864, 2363, 2364]. Наиболее четкие определения этих уровней и наглядное представление о их разграничении приведено в работе [2364]. На основании исследований последних лет к этому можно сделать ряд уточнений: 1) с точки зрения физиологии построения скелета, макроструктуру и текстуру (количество слоев, из которых состоит скелет и его элементы) можно рассматривать как органный уровень формирования скелета; микроструктуру и ультраструктуру - как тканевый; молекулярную структуру - как уровень минералообразования; 2) количество уровней организации скелетной ткани может быть больше (или меньше) пяти в зависимости от числа органических фракций, контролирующих специфичность формирования скелета; 3) при характеристике структуры на любом уровне целесообразно различать собственно структуру (форму) элементарных для этого уровня единиц и их текстуру (взаимное расположение). При образовании скелета биоминерализация проходит в несколько этапов: 1) нуклеация минеральной фазы; 2) первичный рост минеральной фазы; 3) вторичный рост минеральной фазы. Эти этапы, с одной стороны, отражают уровни организации скелетного материала (соответственно молекулярный, ультраструктурный и микроструктурный), а с другой - эволюционные ступени биоминерализации. На каждом из этапов специфичность структуры разных уровней контролировалась соответствующими органическими матрицами, которые включались в скелетную ткань, координируя минеральную фазу скелета и после завершения процесса биоминерализации. Большую, до конца еще не понятую и неоцененную роль органические матрицы играют и в процессах диагенеза и фоссилизации скелетных остатков. Таким образом, скелетная ткань представляет собой сложный минерально-органический комплекс, природный композиционный материал, часто обладающий уникальными свойствами. Кристаллиты разного уровня размерности одеты в чехол органической пленки - матрицы. В этом - главное отличие биоминералов от их аналогов, образованных неорганическим путем и имеющих лишь один уровень иерархии. Степень минерализации и соответственно пропорции органической и неорганической фаз могут быть различны. Различно и количество типов органических матриц и, несомненно, различен их состав. Вопросами состава органических компонентов скелета занимается особая отрасль палеонтологии - палеобиохимия; состав же неорганической компоненты на уровне ее минералогии и главным образом химического элементного состава изучает палеобиогеохимия (см. 2.6). Наглядное, хотя, вероятно, неполное представление о разнообразии органических матриц скелета дало изучение перламутрового слоя моллюсков. Здесь выделяют по меньшей мере три основные органические фракции: 1) "растворимый" накрин - протеин, переходящий в раствор после декальцификации в ЭДТА; 2) "нерастворимая" накрин-протеиновая или гликопротеиновая фракция, переходящая в раствор после обработки кислотами; 3) накроин-органическая фракция (полисахаридная или протеиновая), устойчивая к действию кислот, щелочей, диагенезу и пиролизу. Морфологически накроин - хорошо изученные [1668, 1922] ретикулярные матрицы, обладающие специфичностью на уровне классов моллюсков. "Нерастворимый" накрин, морфологически представленный фибриллярным и глобулярным материалом, слагает межкристаллический и, вероятно, внутрикристаллический (частично) матриксы. Во фракции "растворимого" накрина установлена специфическая последовательность аминокислот в полипептидной цепи протеиновых молекул. Предполагают, что эта фракция ответственна за нуклеацию неорганической фазы скелета, за первичную координацию ее кристаллической решетки [2467]. Именно неоднородность кристаллических и органических компонентов скелетной ткани обусловливает наличие структуры скелетной ткани.
|
К содержанию: «Современная палеонтология»
Смотрите также:
ПАЛЕОНТОЛОГИЯ ПОЗВОНОЧНЫХ геология с основами палеонтологии По следам минувшего
палеоботаника или ботаническая палеонтология... Аллювий прарек Палеовулканология
Древние климаты Палеогеография Палео океанология