|
|
Теломы - элементарные эволюционирующие единицы. Источники параллелизма. Вавилов |
|
Сейчас я должен сделать читателям предупреждение. Его следовало бы сделать, пожалуй, уже в начале главы, но лучше поздно, чем никогда. Дело в том, что мы вступаем в область теоретической ботаники и палеоботаники. Поэтому предлагаю любителям легкого занимательного чтения пролистать книгу дальше и браться за другие книги или журналы. А мы с самыми терпеливыми и любознательными продолжим наше путешествие среди ископаемых растений.
Итак, мы добрались до явлений параллелизма, отметили "коварство" растений, которые не только сами рядятся друг под друга, но заставляют то же проделывать и составляющие их ткани, но ничего не сказали о причинах. Здесь, конечно, легко отделаться магическим заклинанием со ссылкой на внешнюю среду, но попытаемся все же разобраться в механизмах и закономерностях. Для этого воспользуемся испытанными методами.
Когда исследователь сталкивается со сложным явлением, то он прежде всего старается разложить его на составные части. Математик разлагает многочлен, химик выделяет отдельные элементарные реакции, генетик разбирает изменчивость и наследование отдельных признаков. Так же, очевидно, должны поступить и мы, анализируя эволюцию древних растений.
Первым, кто понял это и расчленил растение на элементарные органы (а точнее ‑ на элементарные эволюционирующие единицы), был немецкий палеоботаник Г. Потонье. Он описал массу ископаемых растений, сделал исключительно много в познании пермских и каменноугольных отложений Европы, заложил основы современного учения о накоплении горючих ископаемых, написал массу научно‑популярных статей, издавал журнал и, наконец, сделал огромный вклад в общую морфологию растений. Умер Г. Потонье в 1915 г., когда ему едва перевалило за пятьдесят.
Потонье понял, что лист представляет собой не изначально простое образование, а сложный синтетический орган, результат срастания элементарных структурных единиц. Эта идея была затем развита О. Линье в начале нашего века. Линье различал у растений несколько элементарных единиц. Одни дают начало крупным листьям, типа папоротников, и на них развиваются спорангии; другие представляют выросты первых и всегда лишены спорангиев (это, например, листья плауновидных); третьи дают начало корням. Исходя из своих построений, Линье фактически предсказал открытие псилофитов.
К сожалению, в начале нашего века мало кто из ботаников задумывался над такими проблемами. Поэтому идеи Потонье и Линье прошли почти незамеченными для современников. По выражению одного из современных ботаников, они были "гласом вопиющего в пустыни". Лишь в 30‑х годах немецкий морфолог В. Циммерманн довел эти идеи до логического завершения, достаточно широко распропагандировал их и снабдил свои построения удачной терминологией.
Простые и однообразные органы первых наземных растений ‑ так сказать, элементарные эволюционирующие единицы ‑ он назвал теломами.
Из теломов, претерпевавших различные изменения, образовалось все разнообразие внешних форм у наземных растений. Эти изменения Циммерманн также разложил на элементарные процессы: слияние, изгиб, утолщение, стерилизация и др., которые обеспечивают растению лучшее поглощение света и лучшую защиту спорангиев. Действительно, если спорангий торчит на конце стебля, он максимально подвержен и температурным колебаниям, и различным механическим повреждениям. Загнутая ножка уже дает спорангию преимущество, а еще лучше, если он сидит в пазухе листа под его защитой.
То, что такие элементарные процессы могли происходить у самых разных растений, не вызывает сомнения, и палеоботанические наблюдения это подтверждают. Возьмем, например, плауновидные и членистостебельные. В их эволюции в течение палеозоя и мезозоя наблюдаются одинаковые процессы, приводящие к образованию очень сходных структур. Спорангии, сначала разбросанные по побегу группами, затем собираются в компактные шишечки на концах стеблей. В этих шишках листья преобразуются в специальные образования, защищающие спорангии. Сами спорангии со стебля переходят па эти защитные образования. Сходные процессы проходят у обеих групп и в микроскопической структуре стеблей. Возможно, что плауновидные и членистостебельные связаны общностью происхождения (есть древние растения, которые обнаруживают черты обеих групп), тем более понятно сходство в их эволюции. Таков один из основных источников параллелизма в развитии, но не единственный.
Каждый живой организм, будь то животное или растение, это не простой набор органов, независимо друг от друга выполняющих свои функции. Изменение одного органа вызывает в организации цепную реакцию. Приспособление растений к распространению семян ветром уменьшает размер семян, ведет к изменениям в структуре завязи; развитие новой группы мышц ведет к пропорциональному усилению соответствующего участка нервной системы и т. д. Такие зависимости неисчислимы, и их тем больше, чем более высоко развит организм. Вполне естественно, что реакция разных организмов на одно и то же изменение может оказаться одинаковой, если внутренние зависимости у них также одинаковые. В этом ‑ второй источник параллелизма. К сожалению, здесь трудно привести наглядные примеры, так как для этого надо забираться в терминологические дебри.
Из сказанного не надо делать вывода, что органы вовсе не имеют никакой автономии и являются абсолютно необходимыми. В процессе эволюции они себя чувствуют порой вполне независимо. При этом у растений, даже у самых высокоорганизованных, зависимость между органами значительно меньше, чем у животных. Характерный, хотя и несколько грубый пример: если вы оторвете у собаки хвост, вам не удастся вырастить из него повой собаки, а из веток одного дерева, например тополя, можно вырастить целую рощу нормальных деревьев. Из‑за высокой независимости органов среди растений встречаются удивительные существа, сочетающие архаичные и прогрессивные признаки (подобным дисгармоничным растением является, например, магнолия). Из этого вытекает, что зависимости между органами вряд ли сыграли ведущую роль в параллельной эволюции разных групп растений.
Третий источник параллелизма, по‑видимому, кроется в генетических возможностях живых существ. Генетика сейчас активно популяризируется, с ее терминами все больше и больше знакомится широкая публика. Но на всякий случай приведем два определения. Нам понадобятся в рассуждениях термины "генотип" и "фенотип". Генотип ‑ это вся наследственная информация, которую получает организм в момент его зарождения. В течение жизни она используется не полностью, а лишь в той мере, в какой это допускают условия среды. Результат взаимодействия наследственной информации и среды, т. е. сам выросший организм, называется фенотипом.
В ходе эволюции растения часто упрощают свою структуру. Это доказано, например, для спороносных органов клинолистов каменноугольного периода. В ходе такого упрощения, обычно происходящего через задержку в индивидуальном развитии органа или его частей, былая сложность остается в генотипе. В нем как бы законсервирована возможность последующего, если это станет необходимым, развития сложных структур. И если такая возможность вдруг осуществляется у уже разошедшихся от упрощенного предка ветвей, то нет ничего удивительного, что при этом образуются сходные признаки. Однако мы затрагиваем проблему, которая еще разработана очень мало. Имеющиеся иллюстрации касаются лишь современных растений, а палеоботаника здесь мало чем может помочь.
Генетическую природу имеют и параллелизмы, описанные Н. В. Вавиловым в его ставших классическими гомологических рядах. Хотя он прослеживал параллельную изменчивость на современных культурных растениях, выведенные им закономерности имеют более общее значение. Эти исследования долгое время были вне поля зрения биологов и, к сожалению, вовсе не нашли отклика в палеоботанических обобщениях. Поэтому на гомологических рядах применительно к ископаемым растениям имеет смысл остановиться.
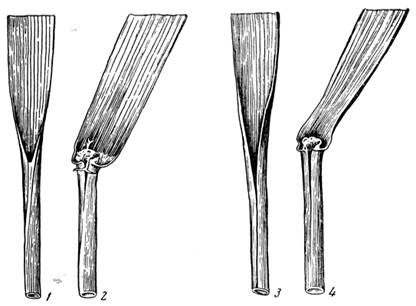
Сущность открытых Вавиловым закономерностей вкратце сводится к следующему. Совершенно очевидно, что чем более родственны друг другу организмы, тем больше у них общих черт. Это исходное положение биологической систематики. В основе каждого вида лежит некий набор обязательных признаков, без которых надо говорить уже о другом виде. Но другие признаки, не являющиеся видовыми, обнаруживают большую или меньшую степень изменчивости. Изучая многочисленные сорта культурных растений, Н. И. Вавилов подметил, что эта изменчивость упорядочена. У близких видов одного рода она идет в почти одинаковых направлениях, причем чем ближе по происхождению виды, тем полнее это сходство. Дело доходит до того, что различие в форме семян (или других органов) у видов, принадлежащих разным родам, становится меньшим, чем у видов одного рода (рис. 37). Получается массовая симуляция органов. То же явление наблюдается и при анализе семейств, порядков и т. д. Но ведь ограниченность изменчивости означает и ограниченность в ассортименте признаков. Растения не могут выйти за пределы определенного круга черт, хотя круг и очень широк.
Параллелизм в строении листьев у пшеницы (1, 2) и ржи (3, 4). По Н. И. Вавилову
Примеры такой ограниченной и повторяющейся изменчивости у систематических групп разного уровня хорошо видны среди ископаемых растений. Когда речь шла о кордаитовых листьях, то фигурировали два их анатомических типа, за которыми стоят разные роды. У одного рода (Rufloria) устьица спрятаны в желобках, а у другого (Cordaites) разбросаны по поверхности листа. По признакам микроструктуры второй род может быть подразделен на несколько подродов. В пределах каждого рода и подрода можно найти листья самого различного внешнего облика: длинные и короткие, узкие и широкие, с закругленной и приостренной верхушкой и т. д. Микроструктура ‑ постоянный устойчивый признак, а внешние черты подвержены значительной, но однообразной изменчивости. Поэтому, классифицируя листья по очертаниям, густоте жилкования и тому подобным признакам без учета микроструктуры, палеоботаники включали в один вид листья, в действительности принадлежащие разным родам.
Прежде чем закончить эти затянувшиеся рассуждения, заметим, что перечисленные пути появления параллелизма не только вероятны, но и доказаны. Однако мы не можем сказать, с каким из них в каждом конкретном случае мы имеем дело. Причина этому ‑ наше почти полное незнание приспособительной ценности самых основных признаков растения. Пожалуй, особенно интересно и непонятно независимое появление очень тонких микроскопических структур в тканях, появление удивительно сходных спор или пыльцы у самых разных групп где‑то в середине или в конце их эволюционного развития. С чем мы здесь имеем дело, пока совершенно непонятно. Конечно, мы очень привыкли относиться к микроструктуре несколько с большим почтением, чем к общей морфологии. Исследователи с легкостью допускают образование одинаковой формы листьев у неродственных растений, а завидев сходство в строении тканей ‑ склонны сразу видеть родство. Однако у нас нет особых оснований считать, что в мастерских природы ювелиры в большей цене, чем плотники, и что их не заставляют одну и ту же работу проделывать заново несколько раз
|
К содержанию: Мейнен: ИЗ ИСТОРИИ РАСТИТЕЛЬНЫХ ДИНАСТИЙ
Смотрите также:
Морфофункциональный анализ. Палеоботаника, палеозоология.
Сложные
случаи отношения формы и функции объединяются понятиями морфофункционального параллелизма
(одна
Второе понятие анализировалось более интенсивно и охватывает разные явления.
Тектология и архитектоника. Изучение многообразия строения...
Речь идет о явлениях,
обозначаемых в литературе как субституция [87], меторизис, гомеозис
В палеоботанике соответствующие исследования широко проводились
еще в прошлом веке и
Метод тройного параллелизма, конкретизируя критерий переходных форм,
указывает, что...